You are browsing environment: FUNGIDB
CAZyme Information: PMAA_091990-t26_1-p1
You are here: Home > Sequence: PMAA_091990-t26_1-p1
Basic Information |
Genomic context |
Full Sequence |
Enzyme annotations |
CAZy signature domains |
CDD domains |
CAZyme hits |
PDB hits |
Swiss-Prot hits |
SignalP and Lipop annotations |
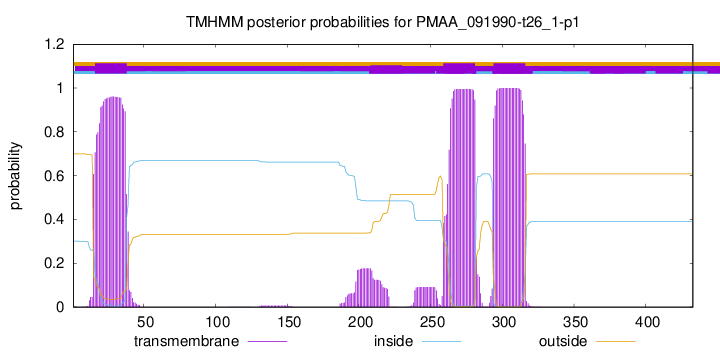
TMHMM annotations
Basic Information help
| Species | Talaromyces marneffei | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lineage | Ascomycota; Eurotiomycetes; ; Trichocomaceae; Talaromyces; Talaromyces marneffei | |||||||||||
| CAZyme ID | PMAA_091990-t26_1-p1 | |||||||||||
| CAZy Family | GT2 | |||||||||||
| CAZyme Description | conserved hypothetical protein | |||||||||||
| CAZyme Property |
|
|||||||||||
| Genome Property |
|
|||||||||||
| Gene Location | ||||||||||||
CAZyme Signature Domains help
| Family | Start | End | Evalue | family coverage |
|---|---|---|---|---|
| GT109 | 25 | 424 | 6.1e-152 | 0.9708029197080292 |
CDD Domains download full data without filtering help
| Cdd ID | Domain | E-Value | qStart | qEnd | sStart | sEnd | Domain Description |
|---|---|---|---|---|---|---|---|
| 409190 | PGAP4-like_fungal | 0.0 | 24 | 395 | 1 | 375 | uncharacterized fungal proteins similar to Post-GPI attachment to proteins factor 4. This subfamily contains uncharacterized fungal proteins with similarity to animal post-GPI attachment to proteins factor 4 (PGAP4), also known as post-GPI attachment to proteins GalNAc transferase 4 or transmembrane protein 246 (TMEM246). PGAP4 has been shown to be a Golgi-resident GPI-GalNAc transferase. Many eukaryotic proteins are anchored to the cell surface through glycolipid glycosylphosphatidylinositol (GPI). GPIs have a conserved core but exhibit diverse N-acetylgalactosamine (GalNAc) modifications. PGAP4 knockout cells lose GPI-GalNAc structures. PGAP4 is most likely involved in the initial steps of GPI-GalNAc biosynthesis. In contrast to other Golgi glycotransferases, it contains three transmembrane domains. Proteins from this subfamily contain the putative catalytic site of PGAP4 and may have similar activities. |
| 409189 | PGAP4-like | 6.35e-57 | 29 | 392 | 1 | 361 | Post-GPI attachment to proteins factor 4 and similar proteins. This family includes post-GPI attachment to proteins factor 4 (PGAP4), also known as post-GPI attachment to proteins GalNAc transferase 4 or transmembrane protein 246 (TMEM246). PGAP4 has been shown to be a Golgi-resident GPI-GalNAc transferase. Many eukaryotic proteins are anchored to the cell surface through glycolipid glycosylphosphatidylinositol (GPI). GPIs have a conserved core but exhibit diverse N-acetylgalactosamine (GalNAc) modifications. PGAP4 knockout cells lose GPI-GalNAc structures. PGAP4 is most likely involved in the initial steps of GPI-GalNAc biosynthesis. In contrast to other Golgi glycotransferases, it contains three transmembrane domains. This family also includes uncharacterized fungal proteins with similarity to PGAP4. |
| 409191 | PGAP4 | 1.75e-23 | 26 | 392 | 4 | 376 | Post-GPI attachment to proteins factor 4. Post-GPI attachment to proteins factor 4 (PGAP4), also known as post-GPI attachment to proteins GalNAc transferase 4 or transmembrane protein 246 (TMEM246), has been shown to be a Golgi-resident GPI-GalNAc transferase. Many eukaryotic proteins are anchored to the cell surface through glycolipid glycosylphosphatidylinositol (GPI). GPIs have a conserved core but exhibit diverse N-acetylgalactosamine (GalNAc) modifications. PGAP4 knockout cells lose GPI-GalNAc structures. PGAP4 is most likely involved in the initial steps of GPI-GalNAc biosynthesis. In contrast to other Golgi glycotransferases (GTs), it contains three transmembrane domains. Structural modeling suggests that PGAP4 adopts a GT-A fold split by an insertion of tandem transmembrane domains. |
| 398374 | Glyco_transf_54 | 5.20e-05 | 97 | 242 | 34 | 186 | N-Acetylglucosaminyltransferase-IV (GnT-IV) conserved region. The complex-type of oligosaccharides are synthesized through elongation by glycosyltransferases after trimming of the precursor oligosaccharides transferred to proteins in the endoplasmic reticulum. N-Acetylglucosaminyltransferases (GnTs) take part in the formation of branches in the biosynthesis of complex-type sugar chains. In vertebrates, six GnTs, designated as GnT-I to -VI, which catalyze the transfer of GlcNAc to the core mannose residues of Asn-linked sugar chains, have been identified. GnT-IV (EC:2.4.1.145) catalyzes the transfer of GlcNAc from UDP-GlcNAc to the GlcNAc1-2Man1-3 arm of core oligosaccharide [Gn2(22)core oligosaccharide] and forms GlcNAc1-4(GlcNAc1-2)Man1-3 structure on the core oligosaccharide (Gn3(2,4,2)core oligosaccharide). In some members the conserved region occupies all but the very for N-terminal, where there is a signal sequence on all members. For other members the conserved region does not occupy the entire protein but is still to the N-terminus of the protein. |
| 407157 | HisKA_7TM | 0.006 | 241 | 323 | 108 | 198 | N-terminal 7TM region of histidine kinase. HisKA_7TM is an N-terminal region consisting of seven transmembrane domains found in Archaea and some bacteria. It is always found associated with histidine kinase. |
CAZyme Hits help
| Hit ID | E-Value | Query Start | Query End | Hit Start | Hit End |
|---|---|---|---|---|---|
| 0.0 | 1 | 433 | 1 | 433 | |
| 3.19e-159 | 24 | 420 | 25 | 422 | |
| 2.10e-157 | 24 | 420 | 25 | 422 | |
| 2.10e-157 | 24 | 420 | 25 | 422 | |
| 5.47e-128 | 111 | 420 | 12 | 323 |
Swiss-Prot Hits help
SignalP and Lipop Annotations help
This protein is predicted as OTHER

| Other | SP_Sec_SPI | CS Position |
|---|---|---|
| 1.000038 | 0.000005 |